
On microbial dormancy and resuscitation
~25% of world's human population contain latent Mycobacterium tuberculosis cells in lung granulomas

When I was researching material for my Systems Thinking post, I came across W Ford Doolittle’s book Darwinizing Gaia. Doolittle was an early critic of the Gaia hypothesis, on the basis that it viewed Earth as an organism upon which has evolved via natural selection. As variations arise during reproduction and the Earth does not reproduce, he found this mechanism untenable. In this latest book, he admits the possibility that ‘persistence’ in addition to differential reproduction could be a selection mechanism.
This got me thinking about the microbial world and the many examples whereby ‘persistence’ operates via a reduced death rate of propagules.
These include production of morphologically recognizable resistant forms:
Endospores formed by Firmicutes..
Akinetes formed by cyanobacteria in the orders Nostocales and Stigonematales. These dense cells can sink to the bottom of aquatic ecosystems and remain dormant until environmental conditions improve.
A significant number of fungal and actinobacterial genera form aerial mycelia that contain conidia which function not only for dispersal but also as metabolically quiescent cells.
Similarly, myxobacteria can form myxospores withing their fruiting bodies, and represent dormant propagules.
Azotobacter species form morphologically-distinguishable cell types called ‘cysts’ that have a reduced metabolic rate and are dessication resistant.
Endospore formation (particularly in Bacillus subtilis) has been studied for many decades as a model of differentiation The volume of literature on conditions leading to dormancy or resuscitation for categories 2-5 is very modest.
Other categories related to dormancy:
1. Viable But Nonculturable (VBNC) cells
2. Persister cells that are not killed by antibiotic treatments
3. Rod-to-coccus transition for several non-mycelial actinobacteria, such as Arthrobacter, Micrococcus and Rhodococcus. [Also note that the actinobacterium Mycobacterium tuberculosis persists in tubercules for years in a dormant state.]
4. Aeonophiles/Anergiobiosis Organisms with incredibly slow metabolic rates that persist in deep subsurface sediments, presumably over geological time scales.
With respect to #3, it feels fair to mention that all rod-shaped cells become smaller as growth rate slows and hence will approximate a spheroid shape. As its doubling times decrease from 120 to 20 minutes, the cell volume of Escherichia coli increases 6-fold. Why should that be? As about 50% of cell mass is protein, growing faster means synthesizing protein faster. Increasing the ribosome content of a cell is a facile way to do that --over that range of doubling times, the cellular ribosome content increases from about 7,000 to 70,000. Increasing the ribosome content of a cell is a facile way to do that and ribosomes take up space. That many ribosomes can only be ‘housed’ in a larger cell volume and hence the small volume of slow growing or dormant cells merely reflect growth rate control of ribosome synthesis.
Measuring dormancy in nature
There are about 250 scientific papers published per year that have dormancy as a topic (over the past 10 years); about one-third of these appear to have an ecological focus. Relatively few actually measure the size of the dormant population – many cite Lennon and Jones (2011) which contains data from the 1990s and 2000s, with a conventional wisdom that most soil bacteria are dormant.
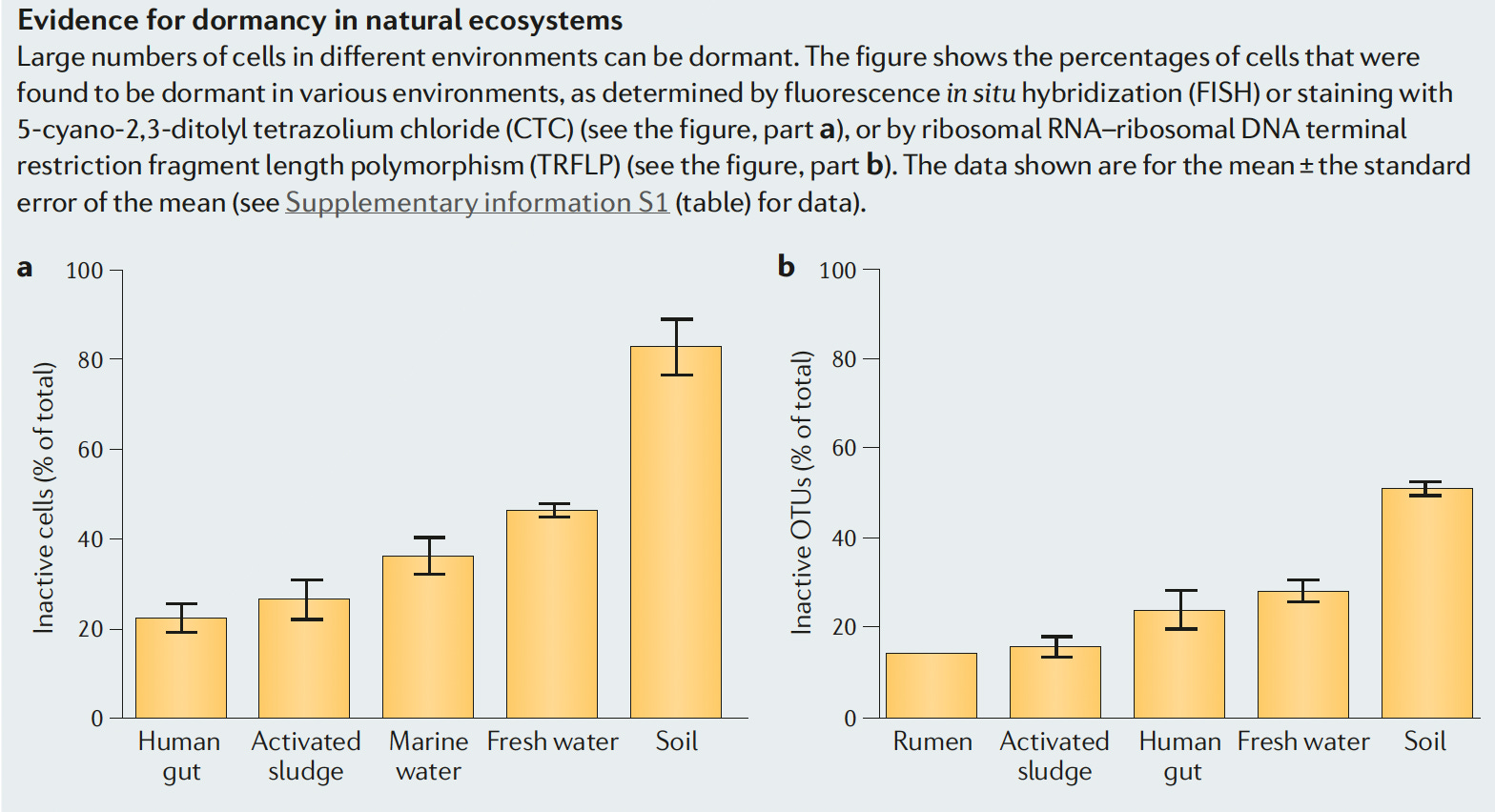
The methods used were the best at the time but certainly have limitations. The most interesting current methods include the use of 18O-water to label nucleic acids (qSIP) and Bioorthogonal Non-canonical Amino Acid Tagging (BONCAT) to label cells actively synthesizing proteins. Harris et al. (2025) applied BONCAT to bulk soil, rhizosphere, plant roots and root nodules and obtained results consistent with conventional wisdom. However, Papp et al. (2018) found that >90% of soil taxa (not cells – important note) incorporated 18O-water into RNA and DNA during incubation periods of 1-8 days. These experiments all “suffer” from a disturbance effect –soils are amended and/or perturbed which is known to stimulate a flush of activity (measured as CO2) evolution. NanoSIMS or Raman-SIP would be an excellent technology with which to combine the advantages of stable isotopes with individual cell measurements.
These latter technologies are much more robust than what was available 15 years ago. Many more analyses of the proportion of metabolically-inactive cells (and the dynamics with which they regain activity upon environmental upshifts) are warranted to move the field from a state of conventional wisdom to a broader analysis of in what habitats and under what circumstances ‘dormancy’ is quantitatively important.
The dynamics of going dormant.
For me, the ‘classic’ paper describing these phenomena was published 30 years ago and summarized a set of work in the Kjelleberg lab on a Vibrio sp.
Kjelleberg, S. et al. How do non-differentiating bacteria adapt to starvation?. Antonie van Leeuwenhoek 63, 333–341 (1993).
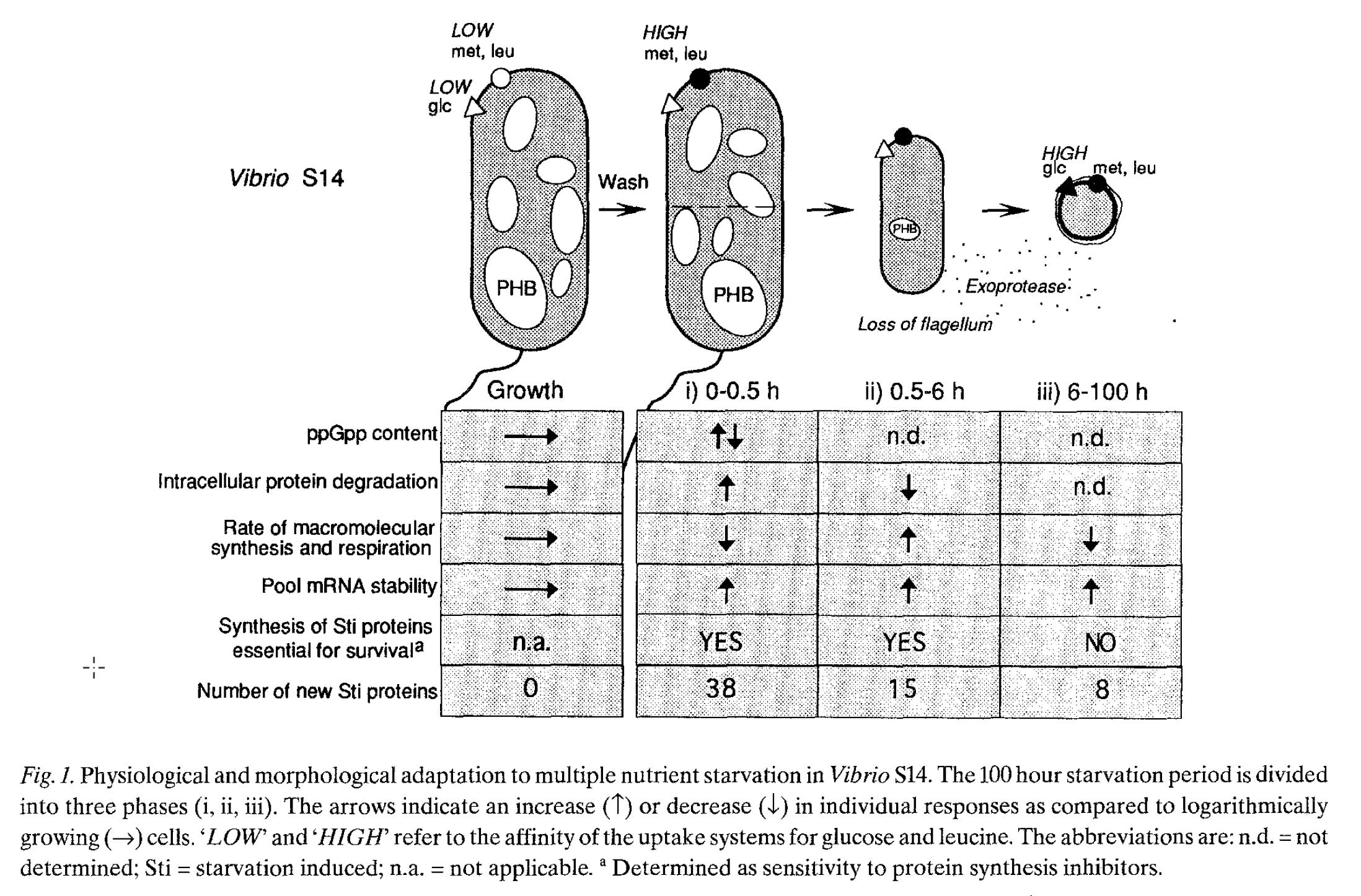
Their work identified several adaptations that have also been identified in a number of less comprehensive studies. Cell size decreases (and becomes spheroid). ppGpp is the stringent control response ‘alarmone’ with pleotropic affects. There are ‘starvation-induced’ proteins; in other studies some of these have been claimed to promote ‘hibernation’ by dimerizing ribosomes and slowing activity of proteins such as RNA polymerase. Note that some of these proteins (in the 0.5-6 h window) cast a wider net for alternative resources (such as an exoprotease or chemotaxis receptors).
Unfortunately, there have been few other studies in the intervening 30 years which have mimicked this analysis using approaches such as transcriptomics. The utility of such experiments would be to determine the extent to which there are general principles that could then be broadly applied or idiosyncratic responses by microbes from particular habitats or evolutionary history.
Yes or no … the game theory of dormancy
Application of the Modern Evolutionary Synthesis posits that ‘dormancy’ is selected because it increases fitness of the organism over evolutionary time scales. But certainly environmental and physiological factors must impact whether fitness improves by going dormant. Some of these include:
How frequently (and regularly) do cells experience starvation, dessication, etc.? For cells in the human GI tract, resources arrive several times a day. Diurnal and tidal rhythms are relatively short term and quite regular. Microbes in a desert soil may experience long periods of aridity, punctuated by brief, intense rainstorms. The aeonophiles apparently persist over millenia without substantial nutrient resources.
Lennon et al. (2021) covered several aspects of theory and how mathematical modeling approaches can be valuable to sort out the environmental and biological dynamics under which transitions to dormancy might have a selective advantage.
Crucial questions include:
How much of a decrease in death rate accrues from the dormant state?
How frequently, regularly and for how long do adverse conditions arise?
How long does resuscitation take after conditions improve?
In a very simple simulation model, Konopka (2000) found that the selection coefficient was most sensitive to the time constant for conversion of dormant cells back to actively growing ones, and the magnitude of the death rate of cells which did not undergo the starvation response
Resuscitation from dormancy
For more than 40 years, microbiologists have been aware that cells can enter a “Viable But Non-Culturable” state and sought means to resuscitate them. James D. Oliver has done extensive work on this topic. Of particular interest are taxa that may be human pathogens, as their survival as VBNC in nature may impact disease transmission or reemergence. Of particular note are intragranulomatous tubercules that are believed to harbor dormant Mycobacterium tuberculosis. Estimates of latent tuberculosis infection range from 20-30% of the world population.
Resuscitation conditions may differ among taxa and include procedures such as temperature shifts. Notably, a number of actinobacteria have been found to generate “Resuscitation promoting factors” and genes have been identified. These appear to be enzymes that enzymes that remodel the peptidoglycan cell wall, and generate fragments that may stimulate other cells to resuscitate. This observation is consistent with Epstein’s scout hypothesis of a few active cells sampling the environment and if there is a growth response, the resulting muropeptide fragments serve as a signal to nearby dormant cells to resuscitate.
As with the dynamics of becoming dormant, transcriptomics is a powerful method to analyze resuscitation. In a study that employed 2H2O as an activity measure in a simulated rain event upon Negev Desert soil, Imminger et al. found that resuscitation was rapid (~30 minutes) and >90% of cells regained activity. Via metatranscriptomics, they were able to identify taxa and specific gene classes that were up-regulated.
Friend-of-the substack Paul Carini is interested in the aridity experienced by microbes in desert soils. He and his colleagues recently published a very interesting paper in which they analyzed the transcriptional responses of an Arthrobacter strain to first dessication, and then rehydration by exposure to *water vapor* rather than water. They identified 6 co-expression modules during these transitions:
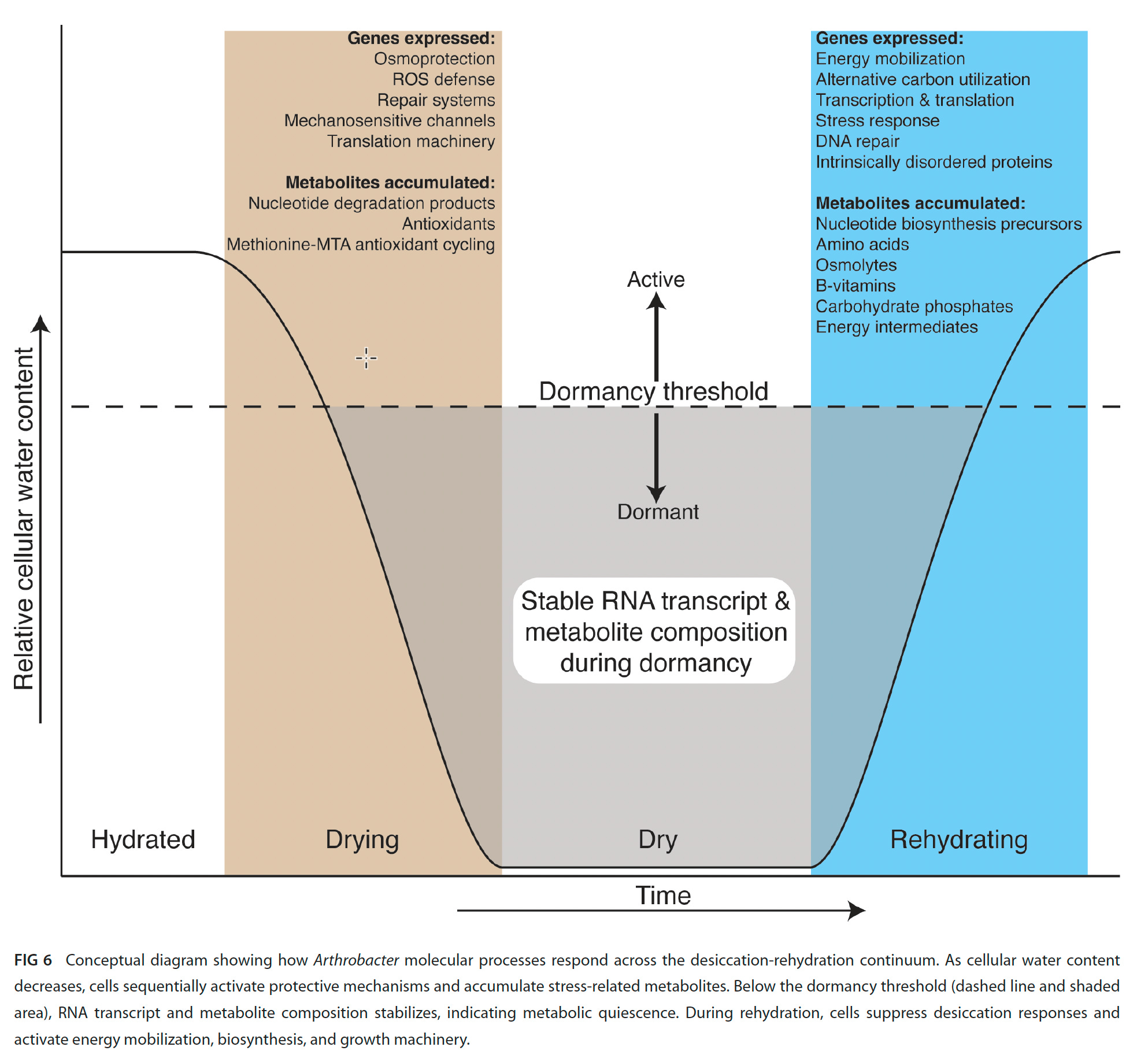
Don Cowan provided a very nice commentary on this article
https://journals.asm.org/doi/10.1128/msystems.00493-25
and some more general comments on the microbial ecology in hyper-arid soils.
In summary, dormancy is an extremely important topic in microbial ecology particularly in certain habitats (unsaturated soils, hyper-arid soils, deep subsurface sediments, and the lesions of those who have been infected by Mycobacterium tuberculosis). However our mechanistic understanding (in the sense discussed here remains very rudimentary, despite the technological capacities to interrogate this phenomenon under ecologically-relevant conditions. I mentioned early in this article that about 80 papers are published per year on ecological aspects of microbial dormancy – that is <1% of all papers published on microbial ecology in 2025.
Today’s Moment of Zen
This is my first posting since returning from a two week trip up the Danube River, starting near Bucharest, Romania and ending near Munich, Germany. This was a great trip in several ways, including my interest in architectural photography. There were a wide variety of architectural styles on offer in cities like Bucharest, Belgrade, Budapest and Vienna. These included Gothic, Renaissance, Baroque, Neo-Classical, Art Noveau, Vienna Secession, Modern, Brutalist, and Deconstruvist. Wow!
Here is a blue-period view of the Hungarian Parliament in Budapest, taken as we sailed upriver toward Bratislava.
