
Doing experiments …

For the last post on my sequence of practical approaches to addressing Big Problems in Microbial Ecology, I am going to focus on modes of experimentation. I recognize that this is currently a minority activity in the discipline. Over the past 10 years, out of 2400 papers published in the Big 5* journals, a Web of Science search indicated only about 400 mentioned experimentation whereas 1300 concerned observations of community composition via DNA sequencing techniques.
The end members of any debate on experimentation are the utility of “real world”/ in situ analyses vs constructed / synthetic communities in the laboratory. Tom Brock defined microbial ecology as the interaction of microbes with their in situ environment. That certainly is the gold standard but can be extremely difficult both operationally and in its ability to refute hypotheses. The use of model systems / “reductionist” approaches is often derided as inferior to analyses of samples taken directly from a habitat, but is it? Or would it be best to not focus on just one approach, but integrate information from different approaches that inform about different scales? Here I discuss the pros and cons of these approaches particularly with a view to identifying principles that may have application not just locally but to other systems.
One of my hard-core principles is that microbial communities are complex systems whose nonlinear dynamics are driven by both external and internal forces that can change in time and space. This makes devising experiments that can unambiguously support or refute hypotheses with in situ samples extremely challenging. Hence, any one sample will differ compositionally from a similar sample at some small distance (cm for soil, tens of cm for the colon, and meters for lakes and oceans. Homogenizing the heterogeneities in these samples only serves to disrupt and likely obscure the interactions occurring at the microenvironment scale.
Analysis of ‘real’ natural samples is certainly warranted in the earliest phases of a research program, when trying to discover what phenomena are important in that habit. For example, the Pace lab made a watershed discovery in 1993 showing that the diversity of a microbial community was much higher than had been thought, via analysis of 16S rDNA genes. However, extracting DNA from a habitat and reporting community composition is in and of itself no longer adding to knowledge in microbial ecology (and yes, there still are papers being published that do only this). The principle has long been established that microbial ecosystems contain a highly diverse set of organisms whose relative abundances generally follow a log normal distribution – the important remaining questions concern how that diversity is maintained.
A disadvantage of authentic natural samples is that analysis is a snapshot of a complex system highly variable in time and space. The experimenter will lack control over a number of environmental factors that may have major impacts. Hence, one is reduced in the end to correlations that may or may not illuminate direct or even indirect causal factors and will have little to say regarding mechanisms.
The appeal of reductionist approaches such as microcosms and synthetic communities are as a means to establish causal principles and mechanisms. In my dream world, there is an interplay between analysis of (complex) phenomena in nature, posing and testing hypotheses under reduced complexity in a better controlled environment and then testing the significance of identified mechanisms back in the natural setting.
This is a variation of John Platt’s 1964 appeal for Strong Inference as a scientific method:
“Strong inference consists of applying the following steps to every problem in science, formally and explicitly and regularly:
1) Devising alternative hypotheses;
2) Devising a crucial experiment (or several of them), with alternative possible outcomes, each of which will, as nearly as possible, exclude one or more of the hypotheses;
3) Carrying out the experiment so as to get a clean result;
1') Recycling the procedure, making sub-hypotheses or sequential hypotheses to refine the possibilities that remain; and so on.”
The ideal of studying microbial ecosystems in situ is most closely approximated with hot spring microbial mats, vertically stratified masses of protoplasm that can be interrogated with microelectrodes at the microscale and which present a ‘natural’ gradient of an important environmental parameter (temperature). The most difficult situation is present in unsaturated soils, with differences in physical and chemical environments at the sub-mm scale. But experiments involving aquatic samples also have their challenges.
Planktonic systems
Doing an experiment (manipulating one or more factors) with an aquatic sample means confining it in some way. ZoBell and Anderson (1936) cautioned that unamended seawater samples incubated in a bottle developed much higher bacterial populations over several days than found in seawater itself and that the effect was inversely proportional to the size of the containers. (However, note that Hammes et al (2010) have disputed this last effect). The explanation has historically been that nutrients adsorb to the bottle surface and bacteria proliferate here.
The consequence for experimental design is that incubation times should be as short as possible (so that isotopic tracers are advantageous for analyzing process rates and introducing a complication for analysis of long-term ecological effects). Larger containers are preferred to smaller ones – for field-oriented studies this has led to the use of microcosms or mesocosms to extend the period of time suitable to monitor conditions akin to the real world.

Microcosm/mesocosm experiments can help eliminate putative mechanisms or estimate field-relevant process rates. They can provide a linkage between authentic field studies and mathematical models that purport to describe system dynamics. But despite the advantages they provide in conducting experiments, in the end they must be supportive to authentic field studies and are best viewed as a heuristic.
Unsaturated soil systems
With its ties to the science of agronomy, soil microbiology has had longstanding experiences with plot experiments in which subsamples can be removed in space or over extended time periods. The soil warming experiment at Harvard Forest is a nice example that has been going on for over 30 years.
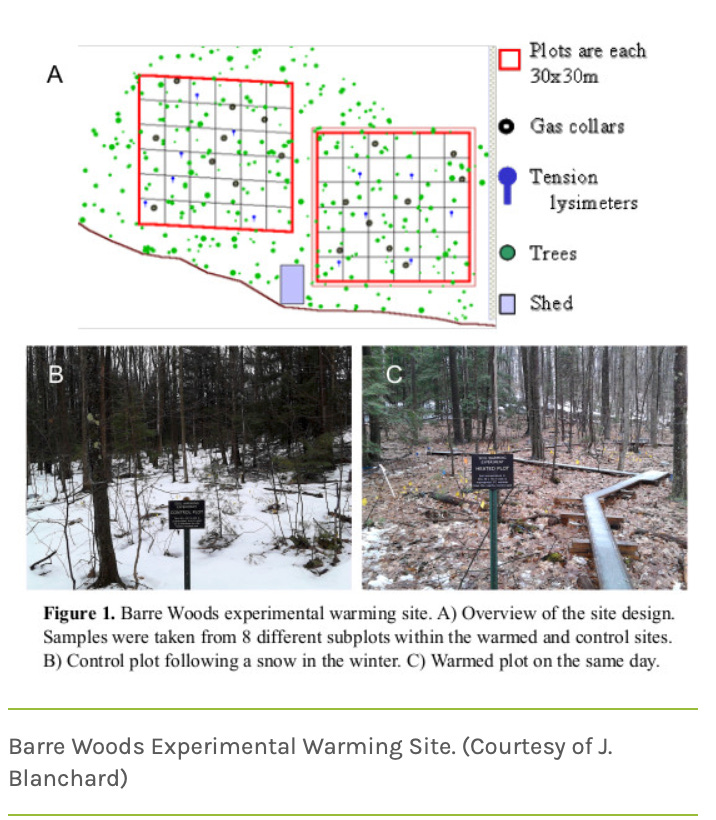
Despite the serious problems of getting at the small-scale spatial heterogeneities present in soil, it is conceivable to carry out experiments at a variety of spatial scales each of which will have strengths and weaknesses that the experimenter should acknowledge when deriving conclusions. As discussed in a previous post, microfluidic devices can mimic the properties present within soil aggregates and get to the microbes’ microenvironment scale. Intact soil cores can be acquired and either incubated in place after experimental amendments (and potentially with vacuum tube lysimeters) or back in more controlled conditions in a laboratory setting, potentially with sampling devices or sensors. (In the case of sediments saturated with water, such cores can be operated as a flow through system, with amendments made to the fluid pumped into the top of the column, and analyses made of samples acquired from sampling ports along the column length).
Let me again emphasize that the greatest knowledge would be obtained from complementary experiments at a variety of spatial scales (from microfluidic devices to plot-scale experiments). Doing so fulfills Platt’s ideal of Strong Inference with experiments at various scales illuminating different aspects of ecosystem dynamics and function. But even with all of this, the interpretation of experimental results will entail extrapolation from the spatial and temporal scales of experimental systems (which of necessity are often quite short) to the much larger and longer scales of the field-scale system.
Reductionist approaches – are they absurd?
I have discussed this a bit with Paul Carini and he characterized these as:
“Reductionist lab conditions. This is where the multidimensionality of the environment is ‘flattened’ to a few dimensions so precise hypotheses can be tested. Gene function studies. Microbial interaction studies, etc. By eliminating the environmental complexity, the experimentalist has a high degree of control.”
These studies certainly are not “real.” But are experiments with natural samples “real”? Any sample is a snapshot of a time and specific place that may not be truly representative of the system dynamics going forward. Per Paul’s comment, studies on model/synthetic microbial communities under better controlled conditions have a better chance of testing hypotheses regarding metabolic interactions, genetic mechanisms and ecological principles governing and structuring communities. Note that perhaps the most deeply held principle in ecology, the competitive exclusion principle, was buttressed by laboratory experiments with 2 Paramecium species.
Jessup et al. (2005) wrote a comprehensive critique and analysis of microbial model systems for ecology. Three key points from their article:
1. The small size and short generation time of microorganisms facilitate replicated experiments across a wide range of spatial and temporal scales
2. The constituents can be well-characterized (this is much more true today than in 2005 due to the prevalence of genome information)
3. Simple systems are used to address fundamental ecological questions. That simplicity may produce more general insights (e.g., competitive exclusion principle) than specific field studies that may be idiosyncratic.
The devil is always in the details and here an important detail is how to choose a suitable model community for one’s research question. This is no easy task. The article by Shetty et al. (2022) is instructive in this regard – they constructed a ‘minimal human gut microbiome’ of 10 species from metagenomic information on community composition and a focus on a particular metabolic function (dietary fiber decomposition). This set allowed ecological processes such as competition, cross-feeding and functional redundancy to occur in the system. Of particular note with respect to Jessup et al.’s point #2 listed above, their analysis of system dynamics entailed transcriptome analysis of the 10 species, tractable because all of their genome sequences were known. Combining this approach with qSIP (quantitative Stable Isotope Probing) employing either 13C or H218O could illustrate primary/secondary nutrient utilization or growth rates respectively.
I hope that what you conclude from this series of 3 posts is that attacking fundamental problems in microbial ecology cannot be accomplished with any one technical approach nor at any one spatial or temporal scale. Yet when I survey the literature that is what I most often see. If I have to point to an exemplar of someone who embraces a broad variety of approaches it would be Kristen DeAngelis at the University of Massachusetts. Her work has entailed both laboratory and field studies, genome analysis and experiments with cultured isolates, experiment and theory. Crucially, she has reached out to collaborators with complementary expertise to advance her work when necessary.
Footnote:
* The Big 5 journals that have microbial ecology as a primary focus are Applied Environmental Microbiology (ecology sections), FEMS Microbiology Ecology, ISME Journal, Environmental Microbiology and Microbial Ecology.
References
Hammes F, Vital M, Egli T.(2010) Critical evaluation of the volumetric "bottle effect" on microbial batch growth. Appl Environ Microbiol. 76:1278-81.
doi: 10.1128/AEM.01914-09.
Jessup, CM et al. (2005) Microbial experimental systems in ecology. Adv Ecolog Res 37: 273-307.
DOI10.1016/S0065-2504(04)37009-1
Platt, JR. 1964. Strong Inference. Science 146: 347
Shetty SA et al. (2022) Inter-species Metabolic Interactions in an In-vitro Minimal Human Gut Microbiome of Core Bacteria. npj Biofilms and Microbiomes 8:21
https://doi.org/10.1038/s41522-022-00275-2
Zobell, C. E. & Anderson, D. Q. (1936) Observations on the multiplication of bacteria in different volumes of stored sea water and the influence of oxygen tension and solid surfaces. Biol. Bull. 71, 324–342.
Your moment of Zen …
I enjoy playing with different post processing techniques, particularly when the ‘straight’ version of an image leaves something to be desired. Here I used a Color Sketch technique for the famous legs of Spanish ham in an Andalusian butcher shop
